Главное меню
Механизмы поддержания долгосрочных ритмов
Механизмы долгосрочного отслеживания времени распространены в природе. Они позволяют организмам подготовиться к изменениям среды задолго до их начала, помогают выживать и успешно размножаться. Цикличность активности гонад, линьки покровов, потребления пищи, массы тела и спячки существует у многих млекопитающих.
Сезонные ритмы точно отлажены, так что детеныши рождаются в наиболее благоприятный период, или например организм мобилизуется перед суровой зимой. Сирийские и сибирские хомяки, овцы, белки, норки, лисы, медведи, олени, обезьяны – наиболее изученные виды, и для каждого из них выявлены специфические наборы сезонных адаптаций.
Указанные представители обитают в условиях севера, но цикличность в репродуктивном периоде необходима и тропическим животным. Так, особи в популяции могут синхронно приступать к размножению, готовиться к сезону дождей или уменьшать потери от хищников (например тропические летучие мыши и лемуры), или же за счет асинхронности снижать конкуренцию среди самцов за право спариваться (например мужские особи азиатского и африканского слона или тропический олень).
Самцы пятнистого оленя, живущего около экватора, имеют схожий тестикулярный ритм секреции, тогда как связанный с ним рост рогов определяется ритмом каждой конкретной особи.
Тропический цирканный ритм сохраняется, даже если животное перевезти в северные широты, передаваясь в исходном варианте во многих поколениях. (Loudon & Curlewis 1988, Lincoln et al. 1996b).
Выделяют два механизма поддержания цирканных притмов у млекопитающих.
Первый – фотопериодизм, это отслеживание годичного изменения длины светового дня и соответствующие изменения в физиологии и поведении.
(Tamarkin et al. 1985).
фотопериодизм осуществляется с помощью фотоиндукции – генетически запрограммированный ответ на смену длинного светового дня на короткий и наоборот.
Этот процесс тесно связан с фоторефрактерностью – изменение в физиологии через недели или месяцы, противоположные начальным. Действуя на подобии отрицательной обратной связи, фоторефрактерность позволяет замкнуть цикл.
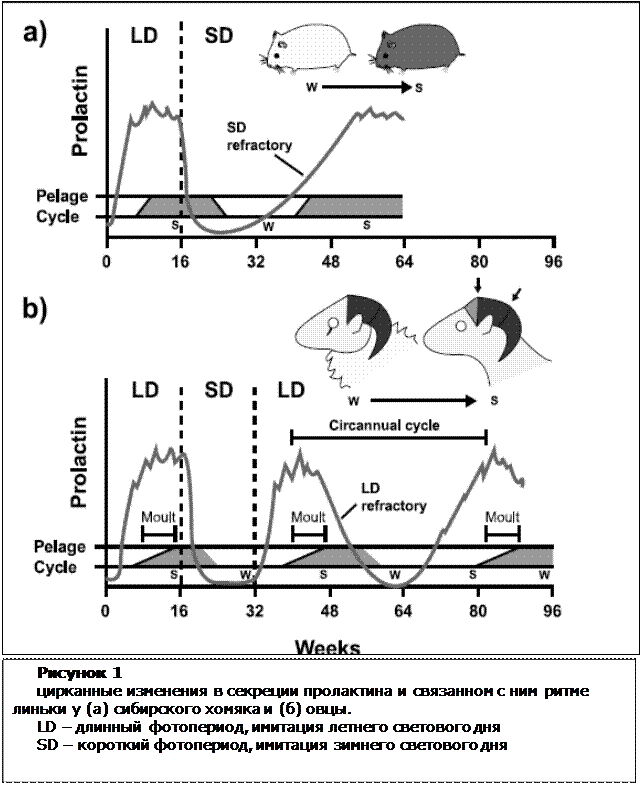
На рисунке 1показан график фотопериод-зависимой секреции пролактина и зависимая от него линька у сибирского хомяка и овец.
Секреция пролактина усиливается на фоне длинного светового периода, запуская физиологические ответы, характерные для лета. Имитирующий зимний период короткий фотопериод, наоборот - угнетает секрецию пролактина.
Сибирские хомяки проявляют фоторефрактерность в ответ на продленный режим короткого светового дня. Так, через 38 недель уровень пролактина достигает летней нормы, также формируется присущая лету окраска агути.
(Bockers et al. 1997, Kuhlmann et al. 2003)
Рефрактерность в данном случае имеет важное биологическое значение: она позволяет животному начать адаптацию к весенне-летнему периоду до выхода из зимней спячки.
Второй механизм представляет собой генерацию непосредственно цирканного ритма. Эта способность была подмечена у многих долгоживущих млекопитающих, независимо от географической широты. Цирканные ритмы продолжают проявлять себя в потомстве, при помещении в зоопарк, несмотря на изменившиеся (выровненные человеком) условия обитания.
(Gwinner 1986, Woodfill et al. 1994)
У некоторых животных (например, зимующей земляной белки, тропических летучих мышей) эндогенный цирканный ритм доминирует, несмотря на несоответствие его фотопериодизму новой среды обитания, тогда как другие животные (например, олени, овцы) подвержены совместному влиянию фотопериодизма и внутреннего цирканного ритма. В последнем случае, фотопериодизм может определять – запускать или нет цирканные ритмы, причем фоторефрактерность при постоянной продолжительности светового дня в таком случае – не отличается от запуска цирканного ритма.
Следовательно, эндогенный регулятор, ответственный за начало фоторефрактерности и генератор цирканных ритмов могут быть единым, наследуемым образованием.
(Prendergast et al. 2002).
Это предполагает наличие единого молекулярного механизма, лежащего в основе генерации цирканных ритмов – подобного тому, что был открыт для циркадианных ритмов.
1 2